

Akphan Behera answered this
Animals need locomotion in the search of food,a better shelter and also for their protection.
- 2
Juno Jayachandran answered this
Animal locomotion, which is the act of self-propulsion by an animal, has many manifestations, including running, swimming, jumping and flying. Animals move for a variety of reasons, such as to find food, a mate, or a suitable microhabitat, and to escape predators. For many animals the ability to move is essential to survival and, as a result, selective pressures have shaped the locomotion methods and mechanisms employed by moving organisms. For example, migratory animals that travel vast distances (such as the Arctic Tern) typically have a locomotion mechanism that costs very little energy per unit distance, whereas non-migratory animals that must frequently move quickly to escape predators (such as frogs) are likely to have costly but very fast locomotion. The study of animal locomotion is typically considered to be a sub-field of biomechanics.
- 0
Anuisri Mandal answered this
Animal locomotion, in ethology, is any of a variety of movements or methods that animals use to move from one place to another. Some modes of locomotion are (initially) self-propelled, e.g., running, swimming, jumping, flying, hopping, soaring and gliding. There are also many animal species that depend on their environment for transportation, a type of mobility called passive locomotion, e.g., sailing (some jellyfish), kiting (spiders) and rolling (some beetles and spiders).
Animals move for a variety of reasons, such as to find food, a mate, a suitable microhabitat, or to escape predators. For many animals, the ability to move is essential for survival and, as a result, natural selection has shaped the locomotion methods and mechanisms used by moving organisms. For example, migratory animals that travel vast distances (such as the Arctic tern) typically have a locomotion mechanism that costs very little energy per unit distance, whereas non-migratory animals that must frequently move quickly to escape predators are likely to have energetically costly, but very fast, locomotion.
Etymology
The term "locomotion" is formed in English from Latin loco "from a place" (ablative of locus "place") + motio "motion, a moving".
Locomotion in different media
Animals move through, or on, four types of environment: aquatic (in or on water), terrestrial (on ground or other surface, including arboreal, or tree-dwelling), fossorial (underground), and aerial (in the air). Many animals—for example semi-aquatic animals, and diving birds—regularly move through more than one type of medium. In some cases, the surface they move on facilitates their method of locomotion.
 Dolphins surfing at Snapper Rocks, Queensland, Australia
Dolphins surfing at Snapper Rocks, Queensland, Australia
In water, staying afloat is possible using buoyancy. If an animal's body is less dense than water, it can stay afloat. This requires little energy to maintain a vertical position, but requires more energy for locomotion in the horizontal plane compared to less buoyant animals. The drag encountered in water is much greater than in air. Morphology is therefore important for efficient locomotion, which is in most cases essential for basic functions such as catching prey. A fusiform, torpedo-like body form is seen in many aquatic animals,[3][4] though the mechanisms they use for locomotion are diverse.
The primary means by which fish and marine mammals generate thrust is by oscillating the body, from side to side in fish, up and down in marine mammals, the resulting wave motion ending at a large tail fin or flippers. Finer control, such as for slow movements, is often achieved with thrust from pectoral fins (or front limbs in marine mammals). Some fish propel themselves without a wave motion of the body, as in the slow-moving seahorses and Gymnotus.[5] Other animals, such as cephalopods, use jet propulsion to travel fast, taking in water then squirting it back out in an explosive burst.[6] Other swimming animals may rely predominantly on their limbs, much as humans do when swimming. Though life on land originated from the seas, terrestrial animals have returned to an aquatic lifestyle on several occasions, such as the fully aquatic cetaceans, now very distinct from their terrestrial ancestors.
Dolphins sometimes ride on the bow waves created by boats or surf on naturally breaking waves.[7]
BenthicBenthic locomotion is movement by animals that live on, in, or near the bottom of aquatic environments. In the sea, many animals walk over the seabed. Echinoderms primarily use their tube feet to move about. The tube feet typically have a tip shaped like a suction pad that can create a vacuum through contraction of muscles. This, along with some stickiness from the secretion of mucus, provides adhesion. Waves of tube feet contractions and relaxations move along the adherent surface and the animal moves slowly along.[8] Some sea urchins also use their spines for benthic locomotion.[9]
Crabs typically walk sideways[10] (a behaviour that gives us the word crabwise). This is because of the articulation of the legs, which makes a sidelong gait more efficient.[11] However, some crabs walk forwards or backwards, including raninids,[12]Libinia emarginata[13] and Mictyris platycheles.[10] Some crabs, notably the Portunidae and Matutidae, are also capable of swimming,[14] the Portunidae especially so as their last pair of walking legs are flattened into swimming paddles.[15]
A stomatopod, Nannosquilla decemspinosa, can escape by rolling itself into a self-propelled wheel and somersault backwards at a speed of 72 rpm. They can travel more than 2 m using this unusual method of locomotion.[16]
Aquatic surface Velella moves by sailing.
Main article: Animal locomotion on the water surface
Velella moves by sailing.
Main article: Animal locomotion on the water surface
Velella, the by-the-wind sailor, is a cnidarian with no means of propulsion other than sailing. A small rigid sail projects into the air and catches the wind. Velella sails always align along the direction of the wind where the sail may act as an aerofoil, so that the animals tend to sail downwind at a small angle to the wind.[17]
While larger animals such as ducks can move on water by floating, some small animals move across it without breaking through the surface. This surface locomotion takes advantage of the surface tension of water. Animals that move in such a way include the water strider. Water striders have legs that are hydrophobic, preventing them from interfering with the structure of water.[18] Another form of locomotion (in which the surface layer is broken) is used by the basilisk lizard.[19]
Aerial Active flight A pair of brimstone butterflies in flight. The female, above, is in fast forward flight with a small angle of attack; the male, below, is twisting his wings sharply upward to gain lift and fly up towards the female.
Main articles: Flight and Flying and gliding animals
A pair of brimstone butterflies in flight. The female, above, is in fast forward flight with a small angle of attack; the male, below, is twisting his wings sharply upward to gain lift and fly up towards the female.
Main articles: Flight and Flying and gliding animals
Gravity is the primary obstacle to flight. Because it is impossible for any organism to have a density as low as that of air, flying animals must generate enough lift to ascend and remain airborne. One way to achieve this is with wings, which when moved through the air generate an upward lift force on the animal's body. Flying animals must be very light to achieve flight, the largest living flying animals being birds of around 20 kilograms.[20] Other structural adaptations of flying animals include reduced and redistributed body weight, fusiform shape and powerful flight muscles;[21][22] there may also be physiological adaptations.[23] Active flight has independently evolved at least four times, in the insects, pterosaurs, birds, and bats. Insects were the first taxon to evolve flight, approximately 400 million years ago (mya),[24] followed by pterosaurs approximately 220 mya,[25] birds approximately 160 mya,[26] then bats about 60 mya.[27][better source needed]
Gliding Main article: Flying and gliding animalsRather than active flight, some (semi-) arboreal animals reduce their rate of falling by gliding. Gliding is heavier-than-air flight without the use of thrust; the term "volplaning" also refers to this mode of flight in animals.[28] This mode of flight involves flying a greater distance horizontally than vertically and therefore can be distinguished from a simple descent like a parachute. Gliding has evolved on more occasions than active flight. There are examples of gliding animals in several major taxonomic classes such as the invertebrates (e.g., gliding ants), reptiles (e.g., banded flying snake), amphibians (e.g., flying frog), mammals (e.g., sugar glider, squirrel glider).
 Flying fish taking off
Flying fish taking off
Some aquatic animals also regularly use gliding, for example, flying fish, octopus and squid. The flights of flying fish are typically around 50 meters (160 ft),[29] though they can use updrafts at the leading edge of waves to cover distances of up to 400 m (1,300 ft).[29][30] To glide upward out of the water, a flying fish moves its tail up to 70 times per second.[31] Several oceanic squid, such as the Pacific flying squid, leap out of the water to escape predators, an adaptation similar to that of flying fish.[32] Smaller squids fly in shoals, and have been observed to cover distances as long as 50 m. Small fins towards the back of the mantle help stabilize the motion of flight. They exit the water by expelling water out of their funnel, indeed some squid have been observed to continue jetting water while airborne providing thrust even after leaving the water. This may make flying squid the only animals with jet-propelled aerial locomotion.[33] The neon flying squid has been observed to glide for distances over 30 m, at speeds of up to 11.2 m/s.[34]
SoaringSoaring birds can maintain flight without wing flapping, using rising air currents. Many gliding birds are able to "lock" their extended wings by means of a specialized tendon.[35] Soaring birds may alternate glides with periods of soaring in rising air. Five principal types of lift are used:[36]thermals, ridge lift, lee waves, convergences and dynamic soaring.
Examples of soaring flight by birds are the use of:
- Thermals and convergences by raptors such as vultures
- Ridge lift by gulls near cliffs
- Wave lift by migrating birds[37]
- Dynamic effects near the surface of the sea by albatrosses
Ballooning is a method of locomotion used by spiders. Certain silk-producing arthropods, mostly small or young spiders, secrete a special light-weight gossamer silk for ballooning, sometimes traveling great distances at high altitude.[38][39]
Terrestrial Main article: Terrestrial locomotion See also: Comparative foot morphology Play media
Pacific leaping blenny Alticus arnoldorum hopping
Play media
Pacific leaping blenny Alticus arnoldorum hopping
 Play media
Pacific leaping blenny climbing up a vertical piece of Plexiglas
Play media
Pacific leaping blenny climbing up a vertical piece of Plexiglas
Forms of locomotion on land include walking, running, hopping or jumping, dragging and crawling or slithering. Here friction and buoyancy are no longer an issue, but a strong skeletal and muscular framework are required in most terrestrial animals for structural support. Each step also requires much energy to overcome inertia, and animals can store elastic potential energy in their tendons to help overcome this. Balance is also required for movement on land. Human infants learn to crawl first before they are able to stand on two feet, which requires good coordination as well as physical development. Humans are bipedal animals, standing on two feet and keeping one on the ground at all times while walking. When running, only one foot is on the ground at any one time at most, and both leave the ground briefly. At higher speeds momentum helps keep the body upright, so more energy can be used in movement.
Jumping Main article: JumpingJumping (saltation) can be distinguished from running, galloping, and other gaits where the entire body is temporarily airborne by the relatively long duration of the aerial phase and high angle of initial launch. Many terrestrial animals use jumping (including hopping or leaping) to escape predators or catch prey—however, relatively few animals use this as a primary mode of locomotion. Those that do include the kangaroo and other macropods, rabbit, hare, jerboa, hopping mouse, and kangaroo rat. Kangaroo rats often leap 2 m[40] and reportedly up to 2.75 m[41] at speeds up to almost 3 m/s (6.7 mph).[42] They can quickly change their direction between jumps.[42] The rapid locomotion of the banner-tailed kangaroo rat may minimize energy cost and predation risk.[43] Its use of a "move-freeze" mode may also make it less conspicuous to nocturnal predators.[43] Frogs are, relative to their size, the best jumpers of all vertebrates.[44] The Australian rocket frog, Litoria nasuta, can leap over 2 metres (6 ft 7 in), more than fifty times its body length.[45]
PeristalsisOther animals move in terrestrial habitats without the aid of legs. Earthworms crawl by a peristalsis, the same rhythmic contractions that propel food through the digestive tract.[46]
SlidingDue to its low coefficient of friction, ice provides the opportunity for other modes of locomotion. Penguins either waddle on their feet or slide on their bellies across the snow, a movement called tobogganing, which conserves energy while moving quickly. Some pinnipeds perform a similar behaviour called sledding.
BrachiationSome animals are specialized for moving on non-horizontal surfaces. One common habitat for such climbing animals is in trees, for example the gibbon is specialized for arboreal movement, traveling rapidly by brachiation. Another case is animals like the snow leopard living on steep rock faces such as are found in mountains. Some light animals are able to climb up smooth sheer surfaces or hang upside down by adhesion. Many insects can do this, though much larger animals such as geckos can also perform similar feats.
Walking and runningSpecies have different numbers of legs resulting in large differences in locomotion.
Modern birds, though classified as tetrapods, usually have only two functional legs, which some (e.g., ostrich, emu, kiwi) use as their primary, Bipedal, mode of locomotion. Few modern mammmalian species are habitual bipeds whose normal method of locomotion is two-legged. These include the macropods, kangaroo rats and mice, springhare,[47]hopping mice, pangolins and homininan apes. Bipedalism is rarely found outside terrestrial animals—though at least two types of octopus walk bipedally on the sea floor using two of their arms, so they can use the remaining arms to camouflage themselves as a mat of algae or floating coconut.[48]
There are no three-legged animals—though some macropods, such as kangaroos, that alternate between resting their weight on their muscular tails and their two hind legs, could be looked at as an example of tripedal locomotion in animals.
Many familiar animals are quadrupedal, walking or running on four legs. A few birds use quadrupedal movement in some circumstances. For example, the shoebill sometimes uses its wings to right itself after lunging at prey.[49] The newly hatched hoatzin bird has claws on its thumb and first finger enabling it to dexterously climb tree branches until its wings are strong enough for sustained flight.[50] These claws are gone by the time the bird reaches adulthood.
A relatively few animals use five limbs for locomotion. Prehensile quadrupeds may use their tail to assist in locomotion and when grazing, the kangaroos and other macropods use their tail to propel themselves forward with the four legs used to maintain balance.
Insects generally walk with six legs—though some insects such as nymphalid butterflies[51] do not use the front legs for walking.
Arachnids have eight legs. Most arachnids lack extensor muscles in the distal joints of their appendages. Spiders and whipscorpions extend their limbs hydraulically using the pressure of their hemolymph.[52]Solifuges and some harvestmen extend their knees by the use of highly elastic thickenings in the joint cuticle.[52]Scorpions, pseudoscorpions and some harvestmen have evolved muscles that extend two leg joints (the femur-patella and patella-tibia joints) at once.[53][54]
The scorpion Hadrurus arizonensis walks by using two groups of legs (left 1, right 2, Left 3, Right 4 and Right 1, Left 2, Right 3, Left 4) in a reciprocating fashion. This alternating tetrapod coordination is used over all walking speeds.[55]
Centipedes and millipedes have many sets of legs that move in metachronal rhythm. Some echinoderms locomote using the many tube feet on the underside of their arms. Although the tube feet resemble suction cups in appearance, the gripping action is a function of adhesive chemicals rather than suction.[56] Other chemicals and relaxation of the ampullae allow for release from the substrate. The tube feet latch on to surfaces and move in a wave, with one arm section attaching to the surface as another releases.[57][58] Some multi-armed, fast-moving starfish such as the sunflower seastar (Pycnopodia helianthoides) pull themselves along with some of their arms while letting others trail behind. Other starfish turn up the tips of their arms while moving, which exposes the sensory tube feet and eyespot to external stimuli.[59] Most starfish cannot move quickly, a typical speed being that of the leather star (Dermasterias imbricata), which can manage just 15 cm (6 in) in a minute.[60] Some burrowing species from the genera Astropecten and Luidia have points rather than suckers on their long tube feet and are capable of much more rapid motion, "gliding" across the ocean floor. The sand star (Luidia foliolata) can travel at a speed of 2.8 m (9 ft 2 in) per minute.[61] Sunflower starfish are quick, efficient hunters, moving at a speed of 1 m/min (3.3 ft/min) using 15,000 tube feet.[62]
Many animals temporarily change the number of legs they use for locomotion in different circumstances. For example, many quadrupedal animals switch to bipedalism to reach low-level browse on trees. The genus of Basiliscus are arboreal lizards that usually use quadrupedalism in the trees. When frightened, they can drop to water below and run across the surface on their hind limbs at about 1.5 m/s for a distance of approximately 4.5 metres (15 ft) before they sink to all fours and swim. They can also sustain themselves on all fours while "water-walking" to increase the distance travelled above the surface by about 1.3 metres.[63] When cockroaches run rapidly, they rear up on their two hind legs like bipedal humans; this allows them to run at speeds up to 50 body lengths/s, equivalent to a "couple hundred miles per hour, if you scale up to the size of humans".[64] When grazing, kangaroos use a form of pentapedalism (four legs plus the tail) but switch to hopping (bipedalism) when they wish to move at a greater speed.
-
 Play media
Play media
Bipedal ostrich
-
 Play media
Play media
Hexapedal stick-insect
-
 Play media
Play media
Octopedal locomotion by a spider
-
 Play media
Play media
Multi-legged millipede
The Moroccan flic-flac spider (Cebrennus rechenbergi) uses a series of rapid, acrobatic flic-flac movements of its legs similar to those used by gymnasts, to actively propel itself off the ground, allowing it to move both down and uphill, even at a 40 percent incline.[65] This behaviour is different than other huntsman spiders, such as Carparachne aureoflava from the Namib Desert, which uses passive cartwheeling as a form of locomotion.[66] The flic-flac spider can reach speeds of up to 2 m/s using forward or back flips to evade threats.[67][68]
SubterraneanSome animals move through solids such as soil by burrowing using peristalsis, as in earthworms,[69] or other methods. In loose solids such as sand some animals, such as the golden mole, marsupial mole, and the pink fairy armadillo, are able to move more rapidly, 'swimming' through the loose substrate. Burrowing animals include moles, ground squirrels, naked mole-rats, tilefish, and mole crickets.
Arboreal locomotion Main article: arboreal locomotion A brachiating gibbon
A brachiating gibbon
Arboreal locomotion is the locomotion of animals in trees. Some animals may only scale trees occasionally, while others are exclusively arboreal. These habitats pose numerous mechanical challenges to animals moving through them, leading to a variety of anatomical, behavioural and ecological consequences as well as variations throughout different species.[70] Furthermore, many of these same principles may be applied to climbing without trees, such as on rock piles or mountains. The earliest known tetrapod with specializations that adapted it for climbing trees, was Suminia, a synapsid of the late Permian, about 260 million years ago.[71] Some invertebrate animals are exclusively arboreal in habitat, for example, the tree snail.
Brachiation (from brachium, Latin for "arm"), is a form of arboreal locomotion in which primates swing from tree limb to tree limb using only their arms. During brachiation, the body is alternately supported under each forelimb. This is the primary means of locomotion for the small gibbons and siamangs of southeast Asia. Some New World monkeys such as spider monkeys and muriquis are "semibrachiators" and move through the trees with a combination of leaping and brachiation. Some New World species also practice suspensory behaviors by using their prehensile tail, which acts as a fifth grasping hand.[72]
EnergeticsAnimal locomotion requires energy to overcome various forces including friction, drag, inertia and gravity, although the influence of these depends on the circumstances. In terrestrial environments, gravity must be overcome whereas the drag of air has little influence. In aqueous environments, friction (or drag) becomes the major energetic challenge with gravity being less of an influence. Remaining in the aqueous environment, animals with natural buoyancy expend little energy to maintain a vertical position in a water column. Others naturally sink, and must spend energy to remain afloat. Drag is also an energetic influence in flight, and the aerodynamically efficient body shapes of flying birds indicate how they have evolved to cope with this. Limbless organisms moving on land must energetically overcome surface friction, however, they do not usually need to expend significant energy to counteract gravity.
Newton's third law of motion is widely used in the study of animal locomotion: if at rest, to move forwards an animal must push something backwards. Terrestrial animals must push the solid ground, swimming and flying animals must push against a fluid (either water or air).[73] The effect of forces during locomotion on the design of the skeletal system is also important, as is the interaction between locomotion and muscle physiology, in determining how the structures and effectors of locomotion enable or limit animal movement. The energetics of locomotion involves the energy expenditure by animals in moving. Energy consumed in locomotion is not available for other efforts, so animals typically have evolved to use the minimum energy possible during movement. However, in the case of certain behaviors, such as locomotion to escape a predator, performance (such as speed or maneuverability) is more crucial, and such movements may be energetically expensive. Furthermore, animals may use energetically expensive methods of locomotion when environmental conditions (such as being within a burrow) preclude other modes.
The most common metric of energy use during locomotion is the net [also termed "incremental"] cost of transport, defined as the amount of energy (e.g., Joules) needed above baseline metabolic rate to move a given distance. For aerobic locomotion, most animals have a nearly constant cost of transport - moving a given distance requires the same caloric expenditure, regardless of speed. This constancy is usually accomplished by changes in gait. The net cost of transport of swimming is lowest, followed by flight, with terrestrial limbed locomotion being the most expensive per unit distance.[20] However, because of the speeds involved, flight requires the most energy per unit time. This does not mean that an animal that normally moves by running would be a more efficient swimmer; however, these comparisons assume an animal is specialized for that form of motion. Another consideration here is body mass—heavier animals, though using more total energy, require less energy per unit mass to move. Physiologists generally measure energy use by the amount of oxygen consumed, or the amount of carbon dioxide produced, in an animal's respiration.[20] In terrestrial animals, the cost of transport is typically measured while they walk or run on a motorized treadmill, either wearing a mask to capture gas exchange or with the entire treadmill enclosed in a metabolic chamber. For small rodents, such as deer mice, the cost of transport has also been measured during voluntary wheel running.[74]
Energetics is important for explaining the evolution of foraging economic decisions in organisms; for example, a study of the African honey bee, A. m. scutellata, has shown that honey bees may trade-off the high sucrose content of viscous nectar for the energetic benefits of warmer, less concentrated nectar, which also reduces their consumption and flight time.[75]
Passive locomotionPassive locomotion in animals is a type of mobility in which the animal depends on their environment for transportation.[1]
Hydrozoans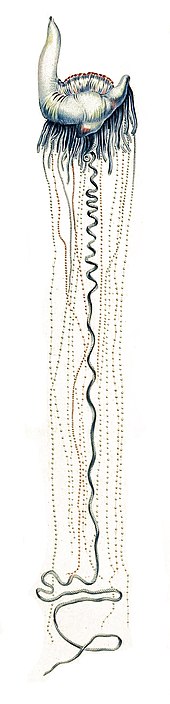 Physalia physalis
Physalia physalis
The Portuguese man o' war (Physalia physalis) lives at the surface of the ocean. The gas-filled bladder, or pneumatophore (sometimes called a "sail"), remains at the surface, while the remainder is submerged. Because the Portuguese man o' war has no means of propulsion, it is moved by a combination of winds, currents, and tides. The sail is equipped with a siphon. In the event of a surface attack, the sail can be deflated, allowing the organism to briefly submerge.[76]
ArachnidsThe wheel spider (Carparachne aureoflava) is a huntsman spider approximately 20 mm in size and native to the Namib Desert of Southern Africa. The spider escapes parasitic pompilid wasps by flipping onto its side and cartwheeling down sand dunes at speeds of up to 44 turns per second.[77][78] If the spider is on a sloped dune, its rolling speed may be 1 metre per second.[79]
A spider (usually limited to individuals of a small species), or spiderling after hatching,[80] climbs as high as it can, stands on raised legs with its abdomen pointed upwards ("tiptoeing"),[81] and then releases several silk threads from its spinnerets into the air. These form a triangle-shaped parachute that carries the spider on updrafts of winds, where even the slightest breeze transports it. The Earth's static electric field may also provide lift in windless conditions.[82]
InsectsThe larva of Cicindela dorsalis, the eastern beach tiger beetle, is notable for its ability to leap into the air, loop its body into a rotating wheel and roll along the sand at a high speed using wind to propel itself. If the wind is strong enough, the larva can cover up to 60 metres (200 ft) in this manner. This remarkable ability may have evolved to help the larva escape predators such as the tiphiid wasp Methocha.[83]
Members of the largest subfamily of cuckoo wasps, Chrysidinae, are generally kleptoparasites, laying their eggs in host nests, where their larvae consume the host egg or larva while it is still young. Chrysidines are distinguished from the members of other subfamilies in that most have flattened or concave lower abdomens and can curl into a defensive ball when attacked by a potential host, a process known as conglobation. Protected by hard chitin in this position, they are expelled from the nest without injury and can search for a less hostile host.
Fleas can jump vertically up to 18 cm and horizontally up to 33 cm,[84] however, although this form of locomotion is initiated by the flea, it has little control of the jump - they always jump in the same direction, with very little variation in the trajectory between individual jumps.[85][86]
CrustaceansAlthough stomatopods typically display the standard locomotion types as seen in true shrimp and lobsters, one species, Nannosquilla decemspinosa, has been observed flipping itself into a crude wheel. The species lives in shallow, sandy areas. At low tides, N. decemspinosa is often stranded by its short rear legs, which are sufficient for locomotion when the body is supported by water, but not on dry land. The mantis shrimp then performs a forward flip in an attempt to roll towards the next tide pool. N. decemspinosa has been observed to roll repeatedly for 2 metres (6.6 ft), but they typically travel less than 1 m (3.3 ft). Again, the animal initiates the movement but has little control during its locomotion.[87]
Animal transportSome animals change location because they are attached to, or reside on, another animal or moving structure. This is arguably more accurately termed "animal transport".
Remoras Some remoras, such as this Echeneis naucrates, may attach themselves to scuba divers.
Some remoras, such as this Echeneis naucrates, may attach themselves to scuba divers.
Remoras are a family (Echeneidae) of ray-finned fish.[88][89] They grow to 30–90 cm (0.98–2.95 ft) long, and their distinctive first dorsal fins take the form of a modified oval, sucker-like organ with slat-like structures that open and close to create suction and take a firm hold against the skin of larger marine animals.[90] By sliding backward, the remora can increase the suction, or it can release itself by swimming forward. Remoras sometimes attach to small boats. They swim well on their own, with a sinuous, or curved, motion. When the remora reaches about 3 cm (1.2 in), the disc is fully formed and the remora can then attach to other animals. The remora's lower jaw projects beyond the upper, and the animal lacks a swim bladder. Some remoras associate primarily with specific host species. They are commonly found attached to sharks, manta rays, whales, turtles, and dugongs. Smaller remoras also fasten onto fish such as tuna and swordfish, and some small remoras travel in the mouths or gills of large manta rays, ocean sunfish, swordfish, and sailfish. The remora benefits by using the host as transport and protection, and also feeds on materials dropped by the host.
Angler fishIn some species of anglerfish, when a male finds a female, he bites into her skin, and releases an enzyme that digests the skin of his mouth and her body, fusing the pair down to the blood-vessel level. The male becomes dependent on the female host for survival by receiving nutrients via their shared circulatory system, and provides sperm to the female in return. After fusing, males increase in volume and become much larger relative to free-living males of the species. They live and remain reproductively functional as long as the female lives, and can take part in multiple spawnings. This extreme sexual dimorphism ensures, when the female is ready to spawn, she has a mate immediately available. Multiple males can be incorporated into a single individual female with up to eight males in some species, though some taxa appear to have a one male per female rule.[91][92]
ParasitesMany endoparasites and ectoparasites, due to their parasitic behaviour, are transported by other animals. For example, tapeworms attach themselves to the inside of the alimentary tracts of other animals and do not locomote within the animal. They do however depend on movement of the host to distribute their eggs.
Other parasites may locomote within, or on, their host, which in turn might be active or stationary. For example, an adult dog flea may crawl about the skin of its sleeping canine host (locomotion), but when the dog awakes and moves, it could be argued the flea is being transported.
Changes between mediaSome animals locomote between different media, e.g., from aquatic to arial. This often requires different modes of locomotion in the different media and may require a distinct transitional locomotor behaviour.
There are a large number of semi-aquatic animals (animals that spend part of their life cycle in water, or generally have part of their anatomy underwater). These represent the major taxons of mammals (e.g., beaver, otter, polar bear), birds (e.g., penguins, ducks), reptiles (e.g., anaconda, bog turtle, marine iguana) and amphibians ( e.g., salamanders, frogs, newts).
FishSome fish use multiple modes of locomotion. Walking fish may swim freely or at other times "walk" along the ocean or river floor, but not on land (e.g., the flying gurnard —which does not actually fly—and batfishes of the Ogcocephalidae family). Amphibious fish, are fish that are able to leave water for extended periods of time. These fish use a range of terrestrial locomotory modes, such as lateral undulation, tripod-like walking (using paired fins and tail), and jumping. Many of these locomotory modes incorporate multiple combinations of pectoral, pelvic and tail fin movement. Examples include eels, mudskippers and the walking catfish. Flying fish can make powerful, self-propelled leaps out of water into air, where their long, wing-like fins enable gliding flight for considerable distances above the water's surface. This uncommon ability is a natural defense mechanism to evade predators. The flights of flying fish are typically around 50 meters,[29] though they can use updrafts at the leading edge of waves to cover distances of up to 400 m (1,300 ft).[29][30] They can travel at speeds of more than 70 km/h (43 mph).[31] Maximum altitude is 6 m (20 ft) above the surface of the sea.[93] Some accounts have them landing on ships' decks.[31][94]
Marine mammals Pacific white-sided dolphins porpoising
Pacific white-sided dolphins porpoising
When swimming, several marine mammals such as dolphins, porpoises and pinnipeds, frequently leap above the water surface whilst maintaining horizontal locomotion. This is done for various reasons. When travelling, jumping can save dolphins and porpoises energy as there is less friction while in the air.[95] This type of travel is known as "porpoising".[95] Other reasons for dolphins and porpoises performing porpoising include orientation, social displays, fighting, non-verbal communication, entertainment and attempting to dislodge parasites.[96] In pinnipeds, two types of porpoising have been identified. "High porpoising" is most often near (within 100 metres) the shore and is often followed by minor course changes; this may help seals get their bearings on beaching or rafting sites. "Low porpoising" is typically observed relatively far (more than 100 metres) from shore and often aborted in favour of anti-predator movements; this may be a way for seals to maximize sub-surface vigilance and thereby reduce their vulnerability to sharks[97]
Some whales raise their (entire) body vertically out of the water in a behaviour known as "breaching".
BirdsSome semi-aquatic birds use terrestrial locomotion, surface swimming, underwater swimming and flying (e.g., ducks, swans). Diving birds also use diving locomotion (e.g., dippers, aulks). Some birds (e.g., ratites) have lost the primary locomotion of flight. The largest of these, ostriches, when being pursued by a predator, have been known to reach speeds over 70 km/h (43 mph),[98] and can maintain a steady speed of 50 km/h (31 mph), which makes the ostrich the world's fastest two-legged animal:[99][100] Ostriches can also locomote by swimming.[101] Penguins either waddle on their feet or slide on their bellies across the snow, a movement called tobogganing, which conserves energy while moving quickly. They also jump with both feet together if they want to move more quickly or cross steep or rocky terrain. To get onto land, penguins sometimes propel themselves upwards at a great speed to leap out the water.
Changes during the life-cycleAn animal's mode of locomotion may change considerably during its life-cycle. Barnacles are exclusively marine and tend to live in shallow and tidal waters. They have two nektonic (active swimming) larval stages, but as adults, they are sessile (non-motile) suspension feeders. Frequently, adults are found attached to moving objects such as whales and ships, and are thereby transported (passive locomotion) around the oceans.
FunctionAnimals locomote for a variety of reasons, such as to find food, a mate, a suitable microhabitat, or to escape predators.
Food procurement Play media
Paddlefish ram suspension-feeding zooplankton in aquarium
Play media
Paddlefish ram suspension-feeding zooplankton in aquarium
Animals use locomotion in a wide variety of ways to procure food. Terrestrial methods include ambush predation, social predation, grazing. Aquatic methods include filterfeeding, grazing, ram feeding, suction feeding, protrusion and pivot feeding. Other methods include parasitism and parasitoidism.
Methods of study Play media
Documentary film, shot at 1200 fps, used to study the locomoton of a cheetah. The end of the video shows the methods used for filming.
Play media
Documentary film, shot at 1200 fps, used to study the locomoton of a cheetah. The end of the video shows the methods used for filming.
A variety of methods and equipment are used to study animal locomotion:
- Treadmills are used to allow animals to walk or run while remaining stationary with respect to external observers. This technique facilitates filming or recordings of physiological information from the animal (e.g., during studies of energetics[102]). Motorized treadmills are also used to measure the endurance capacity (stamina) of animals.[103][104]
- Racetracks lined with photocells or filmed while animals run along them are used to measure acceleration and maximal sprint speed.[105][106]
- Kinematics is the study of the motion of an entire animal or parts of its body. It is typically accomplished by placing visual markers at particular anatomical locations on the animal and then recording video of its movement. The video is often captured from multiple angles, with frame rates exceeding 2000 frames per second when capturing high speed movement. The location of each marker is determined for each video frame, and data from multiple views is integrated to give positions of each point through time. Computers are sometimes used to track the markers, although this task must often be performed manually. The kinematic data can be used to determine fundamental motion attributes such as velocity, acceleration, joint angles, and the sequencing and timing of kinematic events. These fundamental attributes can be used to quantify various higher level attributes, such as the physical abilities of the animal (e.g., its maximum running speed, how steep a slope it can climb), neural control of locomotion, gait, and responses to environmental variation. These, in turn, can aid in formulation of hypotheses about the animal or locomotion in general.
- Force plates are platforms, usually part of a trackway, that can be used to measure the magnitude and direction of forces of an animal's step. When used with kinematics and a sufficiently detailed model of anatomy, inverse dynamics solutions can determine the forces not just at the contact with the ground, but at each joint in the limb.
- Electromyography (EMG) is a method of detecting the electrical activity that occurs when muscles are activated, thus determining which muscles an animal uses for a given movement. This can be accomplished either by surface electrodes (usually in large animals) or implanted electrodes (often wires thinner than a human hair). Furthermore, the intensity of electrical activity can correlate to the level of muscle activity, with greater activity implying (though not definitively showing) greater force.
- Sonomicrometry employs a pair of piezoelectric crystals implanted in a muscle or tendon to continuously measure the length of a muscle or tendon. This is useful because surface kinematics may be inaccurate due to skin movement. Similarly, if an elastic tendon is in series with the muscle, the muscle length may not be accurately reflected by the joint angle.
- Tendon force buckles measure the force produced by a single muscle by measuring the strain of a tendon. After the experiment, the tendon's elastic modulus is determined and used to compute the exact force produced by the muscle. However, this can only be used on muscles with long tendons.
- Particle image velocimetry is used in aquatic and aerial systems to measure the flow of fluid around and past a moving aquatic organism, allowing fluid dynamics calculations to determine pressure gradients, speeds, etc.
- Fluoroscopy allows real-time X-ray video, for precise kinematics of moving bones. Markers opaque to X-rays can allow simultaneous tracking of muscle length.
These methods can be combined. For example, studies frequently combine EMG and kinematics to determine motor pattern, the series of electrical and kinematic events that produce a given movement.
Galleries Swimming in major groups of formerly terrestrial animals Coypu (Rodentia)
Coypu (Rodentia)
 Frog (Anura)
Frog (Anura)
 Sperm whales (Cetacea)
Sperm whales (Cetacea)
 Gentoo penguin (Aves)
Gentoo penguin (Aves)
 Marine iguana (Reptilia)
Flight in major groups
Marine iguana (Reptilia)
Flight in major groups
 Australian Emperor dragonfly (Insecta)
Australian Emperor dragonfly (Insecta)
 Magpie goose (Aves)
Magpie goose (Aves)
 Townsend's big-eared bat (Chiroptera)
Locomotion
Townsend's big-eared bat (Chiroptera)
Locomotion
Animals have evolved an amazing variety of ways to get around. There are animals with no legs; animals with one appendage that serves as a "leg" (snails, clams); animals with two, four, six, or eight legs; animals with dozens of legs; even animals with hundreds of legs. There are animals that move constantly, and animals that stay in one place for their entire adult life. There are animals that swim purposefully and animals that drift wherever the currents take them. Animals slither, crawl, flap, glide, and swim. Some animals spend their entire life underground, whereas others spend almost their entire life in the air. All of these are different modes of animal locomotion.
Locomotion is not the same as movement. All animals move, but not all animals locomote. In ethology , or the study of animal behavior, locomotion is defined as movement that results in progression from one place to another. Animals that spend all or nearly all their entire adult life in one place are called sessile . Animals that move around are called motile.
Locomotion has evolved to enhance the animal's success at finding food, reproducing, escaping predators, or escaping unsuitable habitats. Typically, the animal uses the same mode of locomotion for all these functions, but there are exceptions. For example, a squid normally swims forward or backward by undulating (rhythmically waving) finlike flaps on the sides of its body. However, when startled, the squid expels water through a nozzle and jets backward. Shrimp have a similar behavior. They normally swim using modified appendages called swimerettes. When avoiding a predator, they contract their powerful tail muscles and rapidly move backward through the water. Even some normally sessile animals use crude forms of locomotion to escape predators. Scallops can clap their shells together to produce a sort of jet propulsion. Some cnidarians (such as sea anemones) can break free from their attachment point and then use an undulating motion to swim away from a slow-moving predator.
Principles of LocomotionLocomotion can be passive or active. Each has its advantages and disadvantages. Passive locomotion is the simplest form of animal locomotion. This behavior is exhibited by jellyfish and a few other animals. In this form of locomotion, the environment provides the transportation. The advantage is that no muscular effort is required. The disadvantage of this type of locomotion is that the animal is at the whim of wind and wave. It goes where the current takes it. A somewhat different form of passive locomotion is exhibited by the remora (the name for various species of fish in the family Echeneidae ). Remora attach themselves (harmlessly) to a larger fish or sea turtle and thus go wherever the larger animal goes. However, remora are perfectly capable of swimming on their own.
Most animals exhibit active locomotion at some stage of their life cycle. To move purposefully from place to place, animals must have a means of providing propulsion and a means of controlling their movement. In most cases animals use some sort of muscle tissue attached to a structure to contract and generate the force required to move. This muscle could be attached to a leg bone, causing the animal to jump, as in a frog, or it could contract a chamber, causing a jet of water to propel the animal, as in a squid. The amount, type, and location of contractions are controlled by a nervous system. The nervous system can be as simple as the nerve web in hydra or as complex as the elaborate and highly specialized human nervous system. Nervous system control produces rhythmic movements of the appendages or body that result in locomotion.
Active locomotion can be appendicular or axial. In appendicular locomotion, various appendages such as legs, wings, and flippers interact with the environment by pushing or flapping to produce the propulsive force. Axial locomotion occurs when the animal modifies its body shape to achieve motion. For example, squid contract their large body cavity and forcefully expel water through a nozzle, producing a form of jet propulsion. Eels produce rhythmic ripples down the lengths of their bodies. Leeches stretch out their bodies, extending their anterior ends forward. They then anchor and draw their posterior ends forward by shortening and thickening their bodies.
Whether passive or active locomotion is used, the physical environments occupied by animals fall into four broad categories, each requiring unique forms of locomotion. The four environments are fossorial (underground), terrestrial (on the ground), aerial (in the air, including arboreal , on tree-dwelling), and aquatic (in the water). Each environment has similar restraints on motion: mass or inertia, gravity, and drag. Drag is any force that tends to restrict movement.
In fossorial locomotion, drag is the most important factor restricting forward motion. If the soil is very loose, some animals (insects and lizards) can "swim" through. This form of locomotion is quite rare. Most fossorial animals must burrow or dig tunnels. Some dig as they go, pushing the soil behind them. However, most fossorial animals build permanent tunnels. Once the tunnel is constructed, the mode of locomotion in the tunnel is indistinguishable from terrestrial locomotion.
Animals that spend part of their time in the air (bats, birds, flying insects) need powerful muscles to maintain flight against the force of gravity. Animals that burrow underground or that move about on the surface also require strong muscles to balance the force of gravity. Thus animals that live in aerial, fossorial, or terrestrial environments have evolved strong skeletal systems. Muscles must also overcome inertia to propel the animal forward. The more massive the animal, the more inertia it has.
Many aquatic animals are weightless in water. The buoyancy of the water exactly balances their weight. So muscular effort is not required to maintain their position. However, these animals must still exert muscular effort to initiate motion. Because water has substantial drag, muscular effort is also required to maintain motion. Some animals have negative buoyancy. They sink to the bottom if they stop swimming. Animals with negative buoyancy must expend muscular energy to remain at a given level in the water. An animal with positive buoyancy floats to and rests on or near the surface and must expend muscular energy to remain submerged.
Because the amount of drag due to movement through water is substantial, animals that need to move quickly must have a very streamlined shape. Drag results mainly from the friction of the water as it flows over the surface of the animal. Drag is also caused by water sticking to the surface of the animal. Many fish have evolved a special mucous coating that protects the skin and also reduces friction. The flow of water over the skin of the animal is usually lamellar, which means different layers of the water flow at different speeds relative to the animal. The slowest layer of flow is the one next to the body surface. Moving away from the surface, each layer moves a little faster until the speed of the water flow over the animal is matched at the last layer. Turbulence reduces lamellar flow and increases drag, ultimately limiting the speed of the animal through the water. Dolphins have evolved a gel-like layer just under the skin that tends to absorb turbulence and restores lamellar flow, thus allowing them to swim at a higher speed.
The viscosity of air is much lower than that of water, producing much less drag. However, lamellar flow of air, especially across the wing surfaces, is even more critical. Lift is provided by the shape of the wing. Lift results from air flowing faster across the upper surface than across the lower surface of the wing. Turbulence eliminates lamellar flow and lift is reduced.
Fossorial LocomotionFossorial animals dig burrows, bore into the soil, or construct tunnels. Constructing tunnels or burrows requires that the material be compact and stick together. Semisolid mud or loose sand will not support a burrow. Lizards that "swim" through loose sand or amphibians that swim through mud do not leave tunnels or burrows. While these behaviors could be considered fossorial, they are not discussed here.
Fossorial invertebrates.Burrowing invertebrates have evolved a number of ways to dig through material. Some worms use the contract-anchor-extend method of locomotion. Contraction of the muscles in the rear half of the body pushes the body forward and causes the proboscis to protrude. When the proboscis is fully extended, the worm anchors the proboscis in the soil and pulls the rest of its body forward. This process is repeated, producing a slow and erratic forward motion.
Clams and some other burrowing mollusks use a variation of the contract-anchor-extend method. They extend a muscular "foot" into the soil. Blood is pumped into the foot, causing it to swell and thus forming an anchor. Then the muscle contracts, pulling the clam down into the soil.
Many worms, such as earthworms, use peristaltic locomotion. This form of locomotion is generated by the alternation of longitudinal waves and circular-muscle-contraction waves flowing from the head to the tail. The movement is similar to the contract-anchor-extend method, but each peristaltic wave produces separate anchor points. So several segments of the worm may be moving forward at the same time.
Fossorial vertebrates.Fossorial vertebrates include amphibians, reptiles, and mammals. Locomotion of fossorial amphibians and reptiles is usually axial. Fossorial locomotion of mammals is appendicular. Moles are a good example of fossorial mammals. They have strong, flat forelegs with large, strong claws. Moles dig by extending a foreleg straight ahead in front of the snout and then sweeping it to each side. The loosened soil is pushed against the sidewalls of the burrow. Many rodents dig burrows for nesting but forage above ground. These animals dig by alternately extending their forelegs forward and downward. The loosened soil is pushed backward under the body. The animal may back up through the burrow, pushing the soil out to the surface.
Terrestrial LocomotionThis is the form of locomotion humans use to get around. However, few species use the pure bipedal locomotion of humans. Most animals use four or more legs. Only arthropods and vertebrates have evolved the ability to move rapidly on the ground using legs. Both groups of animals raise their bodies above the ground and use their legs to propel themselves forward. The legs provide both support and propulsion, so the animal must maintain balance as it moves. The sequence and patterns in which the various legs move is determined by the need to maintain balance. More legs create greater stability, but the fastest vertebrates and invertebrates use six or fewer legs.
Walking.Both arthropods and vertebrates use a similar pattern of walking or gait. A foot is planted on the ground and the body is pushed or pulled forward over the foot. The foot remains stationary as the body moves forward. Then the body remains stationary as the foot is lifted and the leg moves forward. For walking and slow running, gaits are generally symmetrical . The footfalls are regularly spaced in time. Fast-moving vertebrates, such as horses, have an asymmetrical but regularly repeating gait.
Insects tend to move their six legs in a simple pattern, lifting and replacing each leg in turn followed by the leg in front of it. Then the legs on the other side are moved. Forward motion always begins with the posterior legs. In slow walking, only one leg is lifted at a time. The limb movements of centipedes and millipedes are similar to those of insects, but with many more legs and simultaneous waves of movement that progress from the posterior end to the anterior end on both sides of the animal.
Four-legged vertebrates must synchronize leg movements to maintain balance. The basic walking pattern of all four-legged vertebrates is left hind leg, left foreleg, right hind leg, and right foreleg. This cycle is then repeated. The faster symmetrical gaits of vertebrates are obtained by overlapping the leg-movement sequences of the left and right sides.
Running.Verterbrates that can run are known as cursorial. They have short, muscular upper legs and thin, elongated lower legs. This adaptation reduces the mass in the lower leg, allowing it to be brought forward more quickly. For slow, steady-running, cursorial vertebrates use a gait known as trotting. All-out running is known as galloping. The gallop is an asymmetrical gait. When galloping, the animal is never supported by more than two legs. Horses at full gallop have all four legs off the ground at the same time during part of the gait. This fact was first demonstrated by Eadweard Muybridge, the American photographer and motion picture pioneer, using highspeed photography involving multiple cameras. His groundbreaking, eleven-volume work, Animal Locomotion, was published in 1899.
Cursorial birds and some lizards use bipedal locomotion. These animals have evolved large feet to increase support. The axis of the body is held perpendicular to the ground. Cursorial birds and lizards have long tails for balance, so that the center of gravity of the animal always falls between its feet. The running gait is, of course, a simple alternation of left and right legs. Lizards begin with four-footed locomotion and switch to bipedal as speed increases.
Hopping.The locomotor pattern of hopping is found in both invertebrates and vertebrates. Invertebrates include a few insects, such as grasshoppers and fleas. Vertebrates include tailless amphibians, kangaroos, rabbits, and a few rodents. All hopping animals have hind legs that are approximately twice as long as the forelegs.
Frogs jump by first flexing their forelegs and tilting their bodies upward. The hind legs are swung out from the sides of the body. When the upper hind leg is perpendicular to the body, the hind leg is forcefully straightened out and the animal is launched upward at a 30° to 45° angle.
Rabbits, kangaroos, and all other mammals move their legs vertically when they jump, instead of horizontally. The hopping gait of rabbits is quadrupedal . A jumping rabbit stretches forward and lands on its forefeet. As the forefeet touch, the back flexes, and the hind end rotates forward and downward. The hind feet touch down next to the forefeet, and a new jump begins. Kangaroos take off and land on their hind feet. The back is not arched and the front legs are used only for balance. All of the muscular effort required for jumping is provided by the powerful hind legs.
Crawling.Invertebrates that crawl use either peristaltic or contract-anchor-extend locomotion. Limbless vertebrates use serpentine, rectilinear, concertina, or sidewinding locomotion. The most common pattern is serpentine locomotion, used by snakes, legless lizards, and a few other species. Rectilinear locomotion is used by most snakes, occasionally by large snakes all the time, and by fossorial limbless vertebrates when burrowing. Concertina and sidewinding locomotion are largely confined to snakes.
Serpentine.In serpentine (snakelike) locomotion, the body moves in a series of curves. In serpentine motion the entire body moves at the same speed. All parts of the body follow the same path as the head. Propulsion is by a lateral thrust in all segments of the body in contact with projections of the surface.
Concertina.Concertina locomotion is used when the surface is too slick for serpentine locomotion. The snake moves its body into a series of tight, wavy loops. These provide more friction on the slick surface. The snake then extends its head forward until the body is nearly straight or begins to slide backward. The snake then presses its head and upper body on the surface, forming a new frictional anchor, and pulls the posterior regions forward.
Sidewinding.Sidewinding locomotion is a specific adaptation for crawling over loose, sandy soils. It may also have the added advantage of reducing contact with hot desert soils. Like serpentine locomotion, the entire body of the snake moves forward continuously in a series of sinuous curves. These curves are sideways to the direction of motion of the snake. The track made by a sidewinding snake is a set of parallel curves roughly perpendicular to the direction of movement. The unique feature of sidewinding is that only two parts of the body touch the ground at any instant. The remainder of the body is held off the ground. To begin, the snake arches the front part of the body forward and forms a loop leaving only the head and the middle of the body in contact with the ground. The snake then moves in a sinuous loop, causing the contact point to move backward along the snake's body as each body segment loops forward. As soon as enough body length is available, the animal forms another loop and begins the next cycle. Each part of the body touches the ground only briefly before it begins to arch forward again.
Rectilinear Locomotion.In snakes, rectilinear motion is completely unlike the other forms of locomotion. The body is held relatively straight and glides forward in a manner similar to the motion of snails. The belly of the snake is covered by rows of wide, overlapping scales. Each scale is attached to two pairs of muscles, both of which are attached at an angle to ribs ahead of and behind the scale. Waves of contraction move from the front of the snake toward the back, lifting and moving each scale forward in turn. Then the scale is pulled rearward, but the edge of the scale digs into the surface, propelling the snake forward.
Aerial and Arboreal LocomotionAnimals have evolved many ways of moving without touching the ground. Aerial locomotion includes gliding, soaring, and true flight. Animals who move through trees are known as arboreal.
Climbing.Each group of arboreal animals has a unique adaptation for climbing. Arthropods weigh little so they show few specialized climbing adaptations. Most arthropods, especially insects, can climb. The heavier vertebrates have many climbing adaptations.
Arboreal frogs and lizards are slender-bodied animals whose climbing gait is essentially the same as their terrestrial gait. The tips of the toes on arboreal frogs are expanded into large, circular disks, which increase the contact area. The digits of arboreal lizards are spread out. On the bottom of each of these spatula-shaped digits are claws and one or two rows of elongated scales. Chameleons have two more specialized adaptations. Their tails are able to grasp objects (prehensile), and their digits have fused into two groups of opposable digits. Chameleons can tightly grasp a thin limb.
Brachiation and leaping.Most arboreal animals must occasionally leap across a gap between trees or branches. The leaping motion is essentially the same as terrestrial leaping, although landing is trickier. Brachiation is using the arms to swing from limb to limb. A few primates have developed highly specialized adaptations for brachiation, although all monkeys brachiate to some extent. Primates that use this form of locomotion have extremely long, powerful arms or forelimbs.
Gliding.In gliding, the animal coasts from a high point to a low point, losing elevation constantly. Gliding animals include amphibians, reptiles, and mammals. The small animals known as flying squirrels demonstrate this behavior. A flying squirrel will climb to near the top of one tree and launch itself into space, gliding to a lower branch on the next tree, then climbing to the top and repeating the process as often as necessary. Gliders have adaptations that allow them to increase the width of their bodies. In the flying squirrels flaps of skin extend from the front limbs to the back. Frogs, snakes, and lizards are able to flatten their bodies. Some gliding lizards have elongated ribs that open like a fan.
Soaring.Soaring is a very different process. Birds who are able to soar are much better gliders than any of the gliding animals. They are able to soar because of their instinctive or learned ability to take advantage of columns of rising air to gain altitude. A vulture will soar in circles in a rising column of air to a high altitude, then glide to the next rising air column. In this way, vultures are able to stay aloft for hours with almost no muscular effort.
True flight.Three living groups of animals possess true flight: insects, birds, and mammals. They can propel themselves upward and forward by flapping their wings. Each of these groups evolved this ability independently of the others. A fourth group, the extinct winged reptiles known as pterosaurs, may have been capable of true flight or only of soaring and gliding. The aerodynamics of flight are basically the same for all flying animals. However, the mechanical details are quite different among the groups. While all three groups propel themselves forward by flapping their wings, many species of birds also include extensive gliding and soaring to conserve energy.
Aquatic LocomotionAnimals that live in aquatic environments exhibit many different forms of locomotion. Some animals crawl or burrow into the bottom of a body of water. Others swim through the water using a variety of different appendages. Still others float freely, following the currents wherever they go. Aquatic organisms range in size from microscopic to the blue whale, the largest animal that has ever lived.
Invertebrates.Aquatic invertebrates swim through the water, crawl along the bottom, or burrow into the bottom. In swimming, muscular activity propels the animal by pushing against the water. On the bottom, muscular activity moves the animal around by interacting with the bottom. Some bottom dwellers simply crawl around on the bottom in a manner exactly like terrestrial locomotion. Others take advantage of the weightless environment to move in ways unique to the water environment.
Aquatic invertebrates have developed two distinct modes of swimming. One mode uses hydraulic propulsion. Jellyfish are a good example of this type of locomotion. They have umbrella-shaped bodies, with the "handle" of the umbrella containing the digestive system. The outer margin of the top of the umbrella, or medusa, is a band of muscles that can contract rapidly. As the muscles contract (just like closing an umbrella) water is expelled forcefully and the jellyfish is propelled along. Scallops use a similar locomotion. They are the best swimmers among bivalves, but at its best, the motion is jerky and poorly controlled. It is used mostly to escape predators. Rapid clapping movements of the two shells create a water jet that propels the scallop.
Cephalopods, such as the squids and octopi, are also mollusks that use water-jet propulsion. Adult cephalopods have lost most of their heavy shell. Many squid are excellent swimmers and can swim forward or backward by undulating flaps along each side of their bodies. All cephalopods are much better swimmers than any other species of mollusk. The mantle of cephalopods encloses a cavity that contains the gills and other internal organs. It also includes, on its bottom surface, a narrow opening called a siphon. When the circular muscles surrounding the cavity simultaneously contract, water is forced through the siphon. This propels the cephalopod in a direction opposite to the direction of the siphon. Thus the siphon also provides directional control.
Fishes.Some fishlike animals use a purely undulatory motion to move themselves. Almost all fish use undulatory movement to some extent and supplement that motion with muscular effort by fins.
An eel swims by undulating its entire body in a series of waves passing from head to tail. This type of movement is called anguilliform (eel-like) locomotion. During steady swimming, several waves simultaneously pass down the body from head to tail. The waves move faster as they approach the animal's tail.
While eels have a body with a fairly blunt anterior and constant diameter for the rest of the length of the body, most fish have a body that tapers at both anterior and posterior ends. For these fish, undulatory motion is not the most efficient. So most fish exhibit carangiform locomotion, in which only the rear half of the body moves back and forth. The fastest swimming fish use this method of locomotion, so it is apparently the most efficient one. In contrast, ostraciiform locomotion uses only the tail fin to sweep back and forth. This is slower and apparently less efficient.
Whales and other cetaceans use undulatory body waves, but the waves move the whale's body up and down instead of from side to side. The elongated tail region of whales produces a form of carangiform locomotion apparently as effective as that of the swiftest fish. Fish, whales, and other aquatic vertebrates have some arrangement of fins distributed around their bodies. They all have a caudal (tail) fin, vertical in fish and horizontal in cetaceans. Aquatic vertebrates also have a large dorsal fin and a pair of large fins (or flippers) on the sides of their bodies close to the front. The caudal fin is the primary means of locomotion. The lateral fins do most of the steering. The dorsal fin or fins provide stability.
Tetrapodal vertebrates.Tetrapodal vertebrates (four-legged vertebrates) that use undulatory locomotion include crocodilians, marine lizards, aquatic salamanders, and larval frogs. However, adult frogs and other tetrapods primarily use appendicular locomotion. Many aquatic tetrapods move primarily by using the hind legs. However, sea turtles, penguins, and fur seals have evolved short hind legs with webbed feet used primarily as rudders. These animals use their powerful forelegs, which have evolved into flippers.
Diving birds, such as cormorants and loons, are propelled by their webbed hind feet. Loons are the best adapted for diving. Their body, head, and neck are elongated and slender; the hind legs have moved far back to the posterior end of the body; the lower legs are short; and the feet are completely webbed.
Frogs and some freshwater turtles have elongated rear legs with enlarged, webbed feet. Other aquatic turtles (such as snapping turtles) are relatively poor swimmers. These turtles walk on the bottom of the lake or stream with limb movements very similar to those used on land except that they can move faster in water than they can on land.
Many mammals have swimming movements identical with their terrestrial limb movements. Most aquatic mammals—such as sea otters, hair seals, and nutria—use their hind legs and frequently their tails for swimming. The feet have some degree of webbing. Fur seals and polar bears swim mainly with forelimbs.
What are the different types of animal locomotion?« Back to Questions ListEthology is the study of animal behavior. Locomotion refers to the movement that causes a progression from one place to another. There are several different types of locomotion exhibited by the animal kingdom. It could either be active or passive. Sessile are animals that spend most of their adult life in one place. Animals that move around are called motile. Corals, sponges are examples of sessile organisms.

The act of flying is called aerial locomotion. Many organisms including birds, insects, bats, flying squirrels, many aquatic species and some amphibians including frog have learnt to fly or glide.
Arboreal locomotion refers to species that live in and move through trees. Leopards are good climbers that can climb up the tree along with their hunted prey to keep them safe from other predators. The challenges of arboreal locomotion include walking on narrow branches, moving up and down the inclines, balancing, swinging with arms from one handhold to another and crossing gaps. Cats, parrots, chameleons, goats, lizards and tree snakes are few examples of arboreal animals.
The movement on water is called aquatic locomotion. This involves swimming or walking on the bottom surface of sea or ocean. Fish, ducks, bacteria, turtles, flat worms, inchworms, leeches etc are organisms that can move through a liquid medium.

Most terrestrial animals move about using cursorial locomotion. Running adaptation of different animals is referred to as cursorial locomotion. Forelimbs and hindlimbs play different roles cursorial four-footed animals. These animals are accustomed to long distance running at high speeds rather than high acceleration over short distances. Cheetahs, wolves, ostriches are known for their cursorial locomotion.
Movement of animals that dig and live underground possess is called fossorial locomotion. Such animals penetrate soil, wood or stone. Many soft bodied invertebrates, moles, earthworms and sea cucumbers are examples of organisms with fossorial locomotion.
Animals using hopping or jumping to move possess saltatorial locomotion. Kangaroos, rabbits and few rodents exhibit saltatorial motion.
- 3